
Ein DNA-Faden besteht aus 140 Basenpaaren. Die DNA liegt spiralförmig im Histon-Komplex als Nukleosom vor. Mehrere Nukleosomen (Oktamer) bilden das perlenschnurartige Hetero-Chromatin. Die Gene werden im entspiralisierten Zustand (Euchromatin) je nach Exons und Introns transkriptiert. Promotorgene, Initiationskodone, Enhancer-Sequenzen und Stoppkodone werden immer in einer Richtung gelesen. Die Methylierung (u.a. am Cytosin) modifiziert die Histon-Bindung und verdichtet das Chromatin. [1] Procain hemmt diese Methylierung bei Krebszellen in vitro. [2]
Man unterscheidet im gesamten Tierreich Wachstums- und Differenzierungsfaktoren (so genannte GDFs). Die wichtigsten sind fibroblast growth factor (FGF), hedgehog- und WNT-Proteine sowie transforming growth factor (TGF-β). 100 verschiedene FGF aktivieren v.a. Tyrosinkinasen (so genannte FGF-Rezeptoren) bei der Mesoderm-Differenzierung, der Angio- und Axonogenese. Hedgehog-Proteine induzieren die Somiten-Bildung, die Neurulation und Knospung der Extremitäten, 15 verschiedene WNT-Proteine regulieren letztere Prozesse und die 30 verschiedenen TGF-β, wozu auch das bone morphogenetic protein (BMP) gehört, jegliche Aufzweigungen in Knospen, Drüsen und Organen. [1]
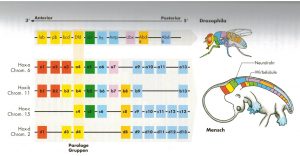
Abb. 1 aus [3]
So genannte Homöobox- bzw. HOX-Gene wurden bei der Fruchtfliege drosophila (Abb.1 aus [3]) entdeckt und bestimmen auch den segmentalen Körperplan von Wirbeltieren. [1,3] So sind die Segmente gegenseitig verknüpft und regulieren sich gegenseitig, d.h. bei Mutation „springt“ das nächste ein. Es verdoppelt sich. [3]
FGF8 wird links des Primitivknotens angereichert. Abhängig von Serotonin (5HT) und Monoaminoxidase (MAO), welche rechts aktiv ist, wird die Neuralplatte induziert. Die Chorda exprimiert weitere Faktoren, welche die Links-Rechts-Determinierung festigen. [1]
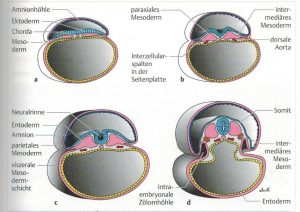
Abb. 2 aus [1]
Im Rahmen der Gastrulation (Einstülpung des Urdarms aus der Blastula) sondern sich mesenchymale Zellen aus dem Ektoderm ab. [4] Als 4. Keimblatt wird die aus der Neurulation hervorgehende Neuralleiste gesehen (Abb.2 aus [1]). Aus ihr entstehen u.a. Hirnnerven, Ganglien, Grenzstrang, Nebennierenmark und Herzseptum. Man unterscheidet mesodermale Zellen, aus den sich zahlreiche Gewebe wie die des muskuloskelattalen und vaskulären Systems entwickeln, vom Mesenchym, was embryonales lockeres Bindegewebe bleibt. Eine Ausnahme stellt das Kopf-Mesenchym dar, was aus der Neuralleiste entsteht. [1]
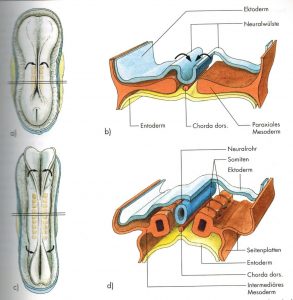
Abb. 3 aus [3]
Die Chorda (axiales Mesoderm) induziert über andere Faktoren die weitere Differenzierung der Keimblätter (v.a. des Ektoderms). Die Segmentierung beginnt in der Mitte (4.Somit) und wird ab 20. Tag von „Urwirbel- bzw. Würfelbildung“ des paraxialen Mesoderms begleitet (Abb.3 aus [3]).
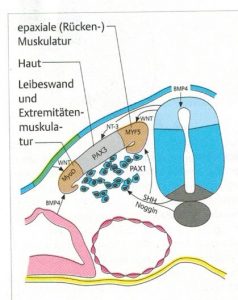
Abb. 4 aus [1]
Die molekulare Steuerung der Differenzierung der Sklerotome übernimmt v.a. das Protein sonic hedgehog (SHH) aus Chorda und Neuralrohr (Bodenplatte). WNT-Proteine aus dem dorsolateralen Neuralrohr induzieren die Myotom-Differenzierung, was BMP hemmend reguliert (Abb.4 aus [1]). Die aus dem paraxialen Mesoderm entstehenden Sklerotome und Dermatomyotome wandern ab 26. Tag aus. Letztere differenzieren sich in Subkutis und Korium sowie Muskeln, welche dorsal das Epimer und ventrolateral das Hypomer bilden (Abb.5 aus [5]).
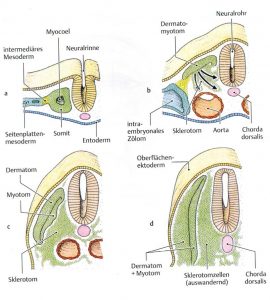
Abb. 5 aus [5]
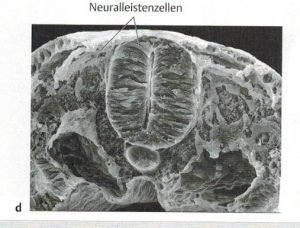
Abb. 6 aus [1]
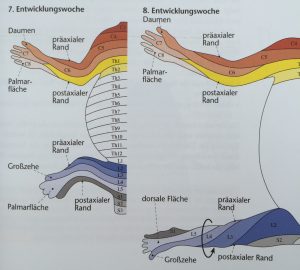
Abb. 7 aus [6]
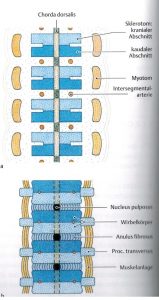
Abb. 8 aus [5]
Sklerotom- und Mesodermanteile aus den Somiten bilden primäre Wirbelkörper, welche zuerst die Chorda und dann das Neuralrohr umwandern. Die Myotome bleiben im primären Nerven-Segment, so dass sich Sklerotome erneut teilen und endgültiges Segment bilden. [3] Mittelpunkt ist und bleibt der Spinalnerv mit den Strukturen im Neuroforamen, welches von Facetten der beiden Wirbelnachbarn umschlossen wird. Durch Bildung der sekundären Segmente (jeweils obere und untere Hälfte des Wirbelkörpers) um eine Arterie entstehen die Bandscheibe, welche einen Chorda-Rest (nucleus pulposus) enthalten. Die Muskeln überbrücken und bewegen die Segmente (Abb.8 aus [5]).
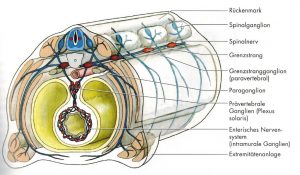
Abb. 9 aus [3]
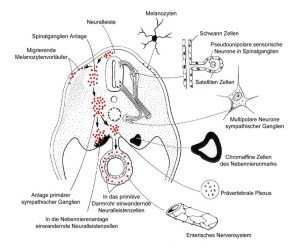
Abb. 10 aus [7]
Das vegetative nervensystem (VNS) differenziert sich vor dem 44. Tag so, dass der Grenzstrang vorwiegend thorakolumbale und der Parasympathikus vorwiegend craniale und pelvine Organe versorgt (Abb.11 aus [3]). Branchiale Organe und Nerven stehen durch die Pharyngeal- bzw. Schlundtaschen bzw. Gill-Bögen [9] segmental in Verbindung:
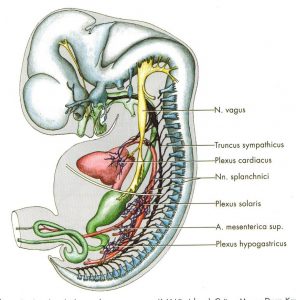
Abb. 11 aus [3]
Der craniale Teil des Parasympathikus nutzt alle Pharyngeal- bzw. Schlundtaschen und -bögen für seine Afferenzen und Efferenzen. In der 5. Woche formieren sich die Kerne der Hirnnerven mit der Formatio retikularis. Alle Kerne (auch Olive, Subst. nigra, Ncll. coerulens, ruber et raphe) entwickeln sich zu eigenen „Steuereinheiten“, (Abb. 12 aus [3]) wobei Ncl. ambiguus und Tractus solitarius eine Sonderrolle in der Poly-Vagal-Theorie nach Porges spielen. [3,9]
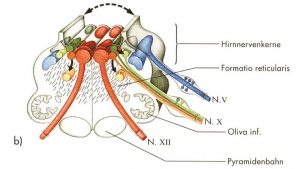
Abb. 12 aus [3]